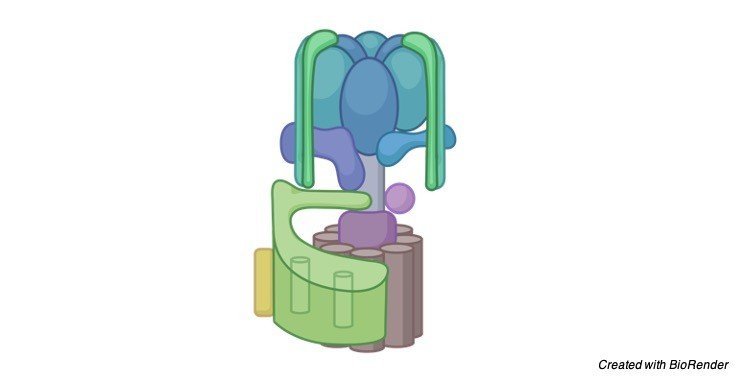
Throughout the long term, proof has gathered in microorganisms and mitochondria that respiratory buildings, including the ATP synthase complex, are coordinated into super-and supramolecular buildings.
Comparable perceptions have been made about photosynthetic buildings in the thylakoid layers of chloroplasts. In the internal layers of mitochondria, dimers of the ATP synthase are shaped by the relationship of their e-subunits in their film areas.
These dimers structure strips, or straight columns, which are confined at the locale of most noteworthy shape along the edges of the cristae.
It is well realized that cardiolipin (diphosphatidylglycerol) is fundamental for the action of a coupled ATP synthase complex.
Cardiolipin atoms have been proposed to tie in the area of a completely trimethylated lysine buildup at position 43 of the c-subunit.
This trimethylated lysine buildup is situated in the headgroup district of the bilayer , and its essence would hamper the limiting of the headgroups of phospholipids, giving destinations to cardiolipin to tie specially.
The ADP–ATP translocase, another plentiful part of the inward layers of mitochondria, additionally contains a trimethyl-lysine buildup, which is likewise in the headgroup area of the layer and would probably have a comparable inclination for restricting cardiolipin atoms.
The suggestion that these trimethyllysine buildups mark cardiolipin-restricting destinations is borne out by atomic elements recreations, and the trimethylated lysine buildup in subunit c is moderated in all vertebrate furthermore, invertebrate ATP synthase edifices that have been inspected, yet not in parasites or microorganisms.
The cardiolipin particles bound to the c8-rings of mitochondrial ATP synthase may balance out the c8- rings to assist them with enduring the rotational force to which they are oppressed.
They may likewise assist with greasing up the pivot of the ring in the film climate. Cardiolipin has been identified additionally in relationship with subunit a.
It is realized that around 75 % of cardiolipin in ox-like heart mitochondria is related with the network pamphlet of the internal layer, and the relationship of cardiolipin with subunit c and the ADP–ATP translocase would contribute to this nearby focus.
In the event that those cardiolipin particles related with ATP synthase buildings were roughly cone-molded, they would in general give the cristae a negative ebb and flow, the inverse to what in particular is noticed.
Consequently, the cardiolipin particles related with ATP synthase are bound to be leveled to the surfaces of the proteins in the layer area of the compound, and their job is probably going to be explicit to the activity of the ATP synthase itself.