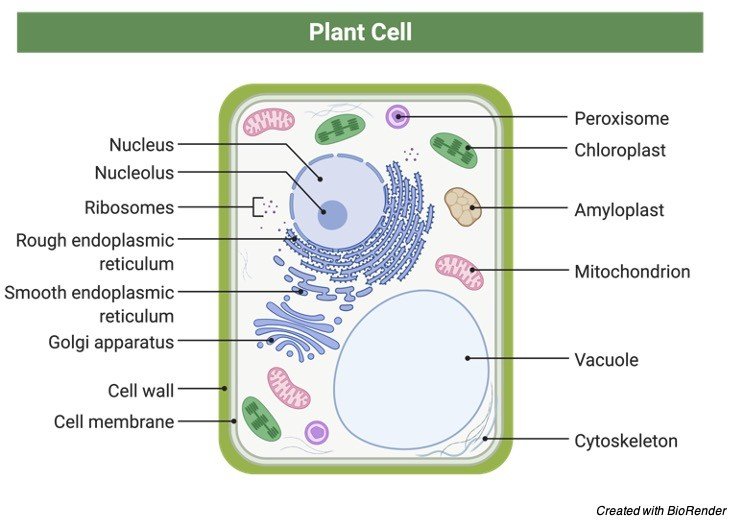
The Golgi apparatus is a membrane-bound sac that modifies and sorts proteins into secretory and transport vesicles. The vesicles are subsequently transported to the plasma membrane and other cell organelles. Although some cells have numerous Golgi apparatuses, most cells have at least one. The apparatus is typically found close to the nucleus.
Endosomes are tubular and vesicular structures that are membrane-bound and situated between the plasma membrane and the Golgi apparatus. By pinching off vesicles or fusing them, they sort and direct vesicular traffic.
Mitochondria are among the most vital organs in the human body. They are the location of a number of chemical reactions that take place during the production of energy packets known as ATP (adenosine triphosphate). There are two membranes that surround each mitochondrion. The outside membrane is smooth, while the inner membrane is folded into cristae-like tubule formations.
Mitochondria are unusual in that they include a small quantity of DNA that contains the genes for the production of certain mitochondrial proteins. Only the mother’s DNA is passed on to the children. Cells with higher activity have more mitochondria, whereas cells with lower activity require fewer energy-producing mitochondria.
A single membrane connects lysosomes, which contain a highly acidic fluid. The fluid contains digestive enzymes that aid in the breakdown of germs and cell waste. They serve a crucial function in the immune system’s cells.
A single membrane also binds peroxisomes. They absorb oxygen and use it to fuel processes that produce hydrogen peroxide by removing hydrogen from different compounds. They are necessary for the cell’s chemical equilibrium to be maintained.
The cytoskeleton is a filamentous network of proteins that are involved in maintaining and changing cell shape as well as cell mobility. The cytoskeleton also creates tracks for cell organelles to move along, which are pushed by contractile proteins on their different surfaces. Inside the cell, it’s like a mini-highway system. The cytoskeleton is made up of three kinds of filaments.
Microfilaments are the cytoskeleton proteins that are the thinnest and most numerous. They are made up of actin, a contractile protein that can be swiftly built and disassembled to meet the demands of the cell or organelle structure.
Intermediate filaments have a somewhat bigger diameter and are present in greater numbers in areas of cells that will be stressed. Filaments will be seen in desmosomes in the skin. These filaments can not be disassembled quickly after they have been assembled.
Microtubules are hollow tubes made out of tubulin, a protein. They are the most stiff and thickest of the filaments. Axons and long dendritic projections of nerve cells include microtubules. They can quickly assemble and disassemble depending on the situation. Microtubules are organised around the centrosome, which contains two centrioles made up of nine sets of fused microtubules. When the centrosome creates the microtubluar spindle fibres required for chromosomal separation during cell division, they are crucial.
Finally, cilia are motile projections on the surface of certain epithelial cells that look like hairs. They feature a 9-set fused microtubule core in the centre. These microtubules cause cilia to move when they interact with a contractile protein. The luminal contents of hollow organs lined by ciliated epithelium are propelled by ciliar motions.