Heterochromatin is the densely packed region of chromatin that impairs the transcription of that particular section of DNA. Euchromatin unlike heterochromatin is lightly packed and genes that are actively transcribed are located in this region.
Chromatin comprises DNA and proteins that bind DNA and stabilize it and it is located in the nucleus in the case of eukaryotes. In its condensed state heterochromatin restricts RNA and DNA polymerases to access the DNA strand and thus, it inhibits both DNA transcription and replication. Heterochromatin is of 2 types facultative and constructive heterochromatin. Heterochromatin constitutes only less than 10% of chromatin in humans.
Heterochromatin Structure
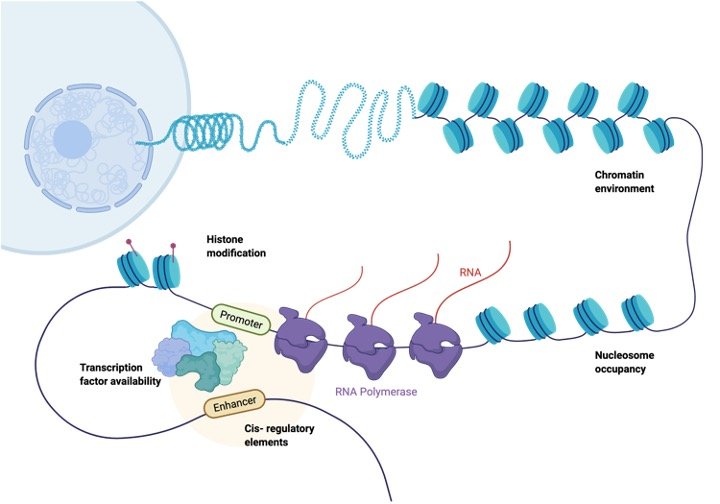
The DNA is present as chromatin that is the complex of proteins and DNA in eukaryotes. The proteins involved in this complex are known as histones that wrap the DNA around them. Approximately 200 bp DNA is wrapped around each histone to form nucleosomes which are sets of 8 histones.
Histones present in nucleosomes are H23, H2A, H4, and H3. Their 2 copies each are present in the nucleosome that form an octamer. Nucleosomes are connected by linker DNA that is strands of DNA connecting both nucleosomes.
This structure of chromatin that is loosely packed is referred to as the beads-on-string model. The beads represent nucleosomes and the string denotes DNA. It depicts how the nucleosomes are connected by DNA comprising chromatin.
Heterochromatin represents the second level of packaging of DNA into a 30-nm condensed fiber that is helically coiled and whose diameter measures 30nm. This type of heterochromatin is more easily visible in G-banding as it is a more tightly condensed form.
The 30 nm fiber structure can be further compacted and condensed into chromosomes and further condensation leads to the formation of metaphase chromosomes.
A double-strand DNA helix is wrapped around an octamer of histones that form nucleosomes to constitute beads on the string structure of euchromatin. It can be further compacted into heterochromatin that comprises 30 nm fiber. Finally, this can be compacted into chromosomes and metaphase chromosomes.
DNA is in the heterochromatin conformation when it is transcriptionally repressed was the polymerases and other regulatory proteins cannot bind to it in this conformation. The linker DNA in euchromatin is available for transcription, while in heterochromatin conformation it is coiled around nucleosomes and is not exposed.
Heterochromatin Types
There are two Types of Heterochromatin: Constitutive and Facultative
Besides the 2 major types of heterochromatin, there are other kinds that intermediate features of facultative and constitutive heterochromatin.
Constitutive heterochromatin stays in heterochromatin configuration and is more stable and often comprises repeated DNA sequences known as satellite DNA. For instance, this kind of heterochromatin is present in telomeres and centromeres that have structural roles.
Facultative heterochromatin is reversible and can change its structure based on the cell cycle stage. It also consists of repeated sequences of DNA called LINE sequences. For example, the Barr body is the inactivated X chromosomes.
Heterochromatin, Cell Cycle, and Gene Expression
DNA packaging level correlates with cell cycle stages. DNA exists in euchromatin form when it either needs to be transcribed or replicated. When the genes in the DNA do not need to be transcribed or replicated then it exists in heterochromatin conformation.
DNA exists in transcriptionally active forms during the interphase stage, while it is present in metaphase chromosome form in mitosis or meiosis stage when the cell is dividing. Thus by regulating DNA conformation or packaging, gene expression can be controlled.
Housekeeping genes are usually present in the euchromatin region as they need to be constitutively expressed as they are involved in metabolic reactions that sustain the cell. The genes that are not actively expressed occur in heterochromatin configuration. This type of gene regulation is achieved by histone tail modifications.
Acetylation or methylation of histone tails can determine the state the DNA exists in. For example, hypoacetylation on the histone tail will repress DNA expression and change the conformation to heterochromatin.
o Sections of DNA that aren’t in use are wrapped tightly around globular proteins called histones.
o Eight histones wrapped in DNA form a nucleosome.
o Nucleosomes, in turn, wrap into coils called solenoids, which wrap into supercoils.
o The entire DNA/protein complex (including a small amount of RNA) is called chromatin.
o The basicity of histones gives them a net positive charge at the normal pH of a cell.
Types of Chromatin
Chromatin is found in two varieties: euchromatin and heterochromatin.
Euchromatin
o Euchromatin is chromatin that can be uncoiled and transcribed.
o Euchromatin is only coiled during nuclear division.
Heterochromatin
o Heterochromatin is a tightly packed form of DNA.
o Its major characteristic is that transcription is limited.
o Some chromatin called constitutive heterochromatin, is permanently coiled.
Chromosomes Structure
o In the nucleus of a human somatic cell, there are 46 double stranded DNA molecules.
o The chromatin associated with each one of these molecules is called a chromosome.
o In human cells, each chromosome possesses a partner that codes for the same traits as itself.
o Two such chromosomes are called homologues.
o Homologous chromosomes are chromosomes in a biological cell that pair (synapse) during meiosis.
o The pair are non-identical chromosomes that both contain information for the same biological features and contain the same genes at the same loci but possibly each have different alleles (that is, different genetic information) at those genes.
o Any cell that contains homologous chromosomes are called diploid.
o Any cell that doesn’t contain homologous chromosomes are called haploid.
o In the nucleus of human cells, there are 46 chromosomes before replication, and 46 chromosomes after replication.
o The duplicates can be referred to separately as sister chromatids.
"Prokaryotic chromosomes exist in the cytoplasm"
o Eukaryotes have multiple pairs of linear chromosomes.
o The gene-coding sequences are interspersed with non-coding regions that may control gene regulation.
o Within a gene sequence, there are exons (coding regions) and introns (non-coding regions that will be removed from the mRNA).
o Eukaryotic chromosomes are contained in a nucleus.
o Prokaryotes have 1 circular chromosome, and may have smaller extrachromasomal DNA in the form of plasmids.
o Prokaryotic DNA does not have the non-coding regions or introns.
o Bacteria do NOT have histones, but archaea have histone-like structures associated with their chromosomes.
o Prokaryotic chromosomes exist in the cytoplasm.
o Reality about Bacterial Chromosomes: Although many have only the 1 circular chromosome, there are many exceptions to this rule.
We all know that our body is made up of millions of cells, each cell makes up the structural and functional unit of life. Each cell consists of many cellular organelles such as nucleus, mitochondria, lysosome, etc.
Chromosomes are one of the important components which are present in the nucleus of the cell and contains an organized package of DNA, which is very much important in carrying out the body’s metabolic and enzymatic activities and also has a vital i role in carrying out the hereditary characters.
Chromatin is one of the complexes of the DNA and proteins which forms the chromosomes. Chromatin generally occurs in two forms, namely euchromatin and heterochromatin.
Euchromatin is less condensed and can be transcribed, whereas the heterochromatin is highly condensed and is not typically transcribed.
What is Heterochromatin?
Heterochromatin is a condensed or a tightly packed form of DNA, which comes in various forms. These forms stay in a continuous manner in between the two ends of the constitutive heterochromatin and facultative heterochromatin.
These two play an important role in expressing the genes. As it tightly packed, it was said to be inaccessible to the polymerases and therefore it cannot be transcribed.
If much of this DNA, is transcribed. It is continuously turned over the RNA induced transcriptional silencing, recent studies suggest that electron microscopic staining reveal that the dense packing is not due to the chromatin.
Constitutive heterochromatin affects the genes near itself. It occurs in the repetitive manner and it performs structural functions such as centromeres and telomeres.
It acts as an attractor along with expressing the genes and also in repressing signals.
Facultative heterochromatin is due to the result of genes which are silenced through an activity of the histone deacetylation or Piwi-interacting RNA, which is commonly called as PiRNA through RNAi.
This does not occur in a repetitive manner and it also shares the compact structure of the consecutive heterochromatin.
Under specific developmental or environmental signaling of cues, it can sometimes lose its condensed structure and it becomes transcriptionally active.
Heterochromatin is also associated with the di and tri methylation of H3K9 in some certain portions of the genomes. Here H3K9me3 related methyl transferases appears to have pivotal role in modifying the heterochromatin during the lineage commitment which occurs at the onset of organogenesis and it also maintains the lineage fidelity.
Structure of Heterochromatin
Chromatin usually occurs in two varieties such as euchromatin and heterochromatin. These two forms are distinguished cytologically by assuming how intensely they are getting stained; The euchromatin is stained lightly, whereas the heterochromatin is stained intensely such that it looks dark in color, which indicates that it is a tight packaging.
Heterochromatin is usually located at the end of the nucleus. Inspite of this early dichromia, recent evidences in both animals and plants suggest that there were more than two different states of heterochromatin.
But it has also been said that it exists in four or five states which are marked with different combinations in the epigenetic marks.
Heterochromatin usually has its genetically inactive satellite sequences and many of the genes which represses the various extents.
Even though some of them are not able to express their genes in euchromatin at any of the time. Where as both centromeres and telomeres are heterochromatic, as such as how it is present in the Barr body to the second inactivated X-chromosome n the female.
Function of Heterochromatin
Heterochromatin is usually associated with several functions which forms the regulation of gene to protect the integrity of chromosome.
Some roles of heterochromatin help in attributing the dense packaging of DNA, which helps in making it less accessible to the protein factors to bind DNA or its other associating factors.
Some regions of chromatin are very densely packaged along with the fibers which displays a condition compared to that of the chromosomes at its mitosis stage.
Heterochromatin is usually inherited by cloning, During the process of cell division the two daughter cells are produced which contains heterochromatin, in the same regions of DNA which results in epigenetic inheritance.
Variations in the heterochromatin encroaches on the adjacent genes or recedes from the genes to the extreme of the domains.
Transcribable material can be expressed by positioning at the boundary domains. This gives rise to the levels of expression that are varying in the cells from one cell to the other.
Constitutive Heterochromatin
Usually, all cells in many of the species has the same regions of the DNA as their packaging sites, especially in a constitutive heterochromatin and as a result any genes which is present within the constitutive heterochromatin in all the cells are poorly expressed.
In most of the organisms this constitutive heterochromatin occurs around the region of chromosomal centromere and near the telomeres.
Facultative Heterochromatin
The regions of the DNA which are packaged in the facultative heterochromatins are not consistent in all the cell type within the pieces, and hence the sequence on one cell is packed in facultative heterochromatin, which means that the genes are expressed poorly so that it can packed in a euchromatin of another cell.
However, the formation of this facultative heterochromatin is regulated often and it is associated with the morphogenesis or differentiation.
Yeast Heterochromatin
One of the species of yeast, such as Saccharomyces cerevisiae, which is also commonly called as budding yeast is considered as one of the eukaryotes and it also has a definite structure of heterochromatin.
Most of its genome are characterized as euchromatin where the regions of the DNA are transcribed very poorly, this type of loci are called as silent mating type loci, the rDNA, sub telomeric regions.
Where as the fission yeast uses another mechanism for the formation of heterochromatin in its centromeres.