Their principal work is combination and capacity of fats and oils. They contain something like 50 distinct enzymes, which are engaged with an assortment of biochemical pathways in various sorts of cells.
They complete oxidation responses prompting the combination of hydrogen peroxide and in light of the fact that it is unsafe to the cell, peroxisomes additionally contain the enzyme catalase, which deteriorates hydrogen peroxide either by changing it over to water or by utilizing it to oxidize another natural compound.
An assortment of substrates are separated by such oxidative responses in peroxisomes, including uric corrosive, amino acids, and unsaturated fats. The oxidation of unsaturated fats is an especially significant model since it gives a significant wellspring of metabolic energy.
In creature cells, unsaturated fats are oxidized in both peroxisomes and mitochondria, however in yeasts and plants unsaturated fat oxidation is confined to peroxisomes.
As well as giving a compartment to oxidation responses, peroxisomes are engaged with lipid biosynthesis. In creature cells, cholesterol and dolichol are integrated in peroxisomes just as in the ER. In the liver, peroxisomes are additionally associated with the amalgamation of bile acids, which are gotten from cholesterol.
What’s more, peroxisomes contain enzymes needed for the union of plasmalogens—a group of phospholipids wherein one of the hydrocarbon ties is joined to glycerol by an ether bond as opposed to an ester bond.
Plasmalogens are significant film segments in certain tissues, especially heart and mind, despite the fact that they are missing in others.
Peroxisomes assume two especially significant parts in plants. To start with, peroxisomes in seeds are liable for the change of put away unsaturated fats to sugars, which is basic to giving energy and crude materials to development of the growing plant.
This happens through a progression of responses named the glyoxylate cycle, which is a variation of the citrus extract cycle. The peroxisomes in which this happens are now and then called glyoxysomes.
Second, peroxisomes in leaves are associated with photorespiration, which serves to use a side item shaped during photosynthesis. CO2 is changed over to starches during photosynthesis through a progression of responses called the Calvin cycle.
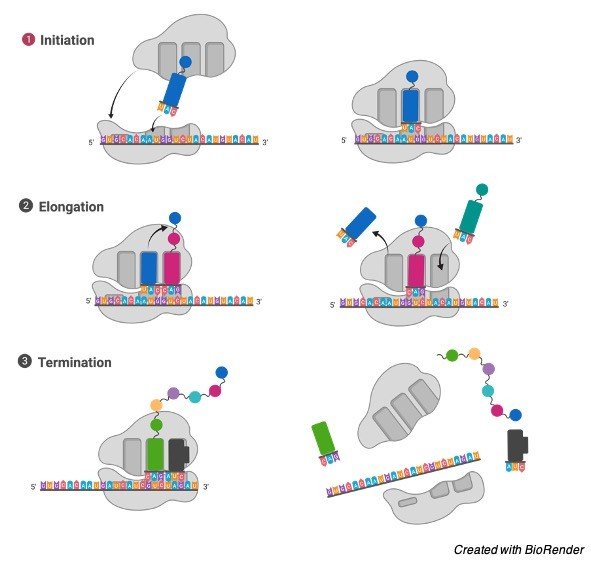
Peroxisome gathering: Proteins bound for peroxisomes are deciphered on free cytosolic ribosomes and afterward shipped into peroxisomes as finished polypeptide chains.
Phospholipids are likewise imported to peroxisomes, by means of phospholipid move proteins, from their significant site of union in the ER.
The import of proteins and phospholipids results in peroxisome development, and new peroxisomes are then framed by division of old ones.
It is a solitary layer bound vesicle with a breadth of about 0.5 mm having a gathered wellspring of something like three oxidative enzymes in liver cells: D-amino corrosive oxidize, urate oxidize and catalase.